Article preview View full access options
Nature | Article
A new antibiotic kills pathogens without detectable resistance
- Journal name:
- Nature
- Year published:
- DOI:
- doi:10.1038/nature14098
- Received
- Accepted
- Published online
Abstract
Antibiotic resistance is spreading faster than the introduction of new compounds into clinical practice, causing a public health crisis. Most antibiotics were produced by screening soil microorganisms, but this limited resource of cultivable bacteria was overmined by the 1960s. Synthetic approaches to produce antibiotics have been unable to replace this platform. Uncultured bacteria make up approximately 99% of all species in external environments, and are an untapped source of new antibiotics. We developed several methods to grow uncultured organisms by cultivation in situ or by using specific growth factors. Here we report a new antibiotic that we term teixobactin, discovered in a screen of uncultured bacteria. Teixobactin inhibits cell wall synthesis by binding to a highly conserved motif of lipid II (precursor of peptidoglycan) and lipid III (precursor of cell wall teichoic acid). We did not obtain any mutants of Staphylococcus aureus or Mycobacterium tuberculosis resistant to teixobactin. The properties of this compound suggest a path towards developing antibiotics that are likely to avoid development of resistance.
At a glance
Figures
-
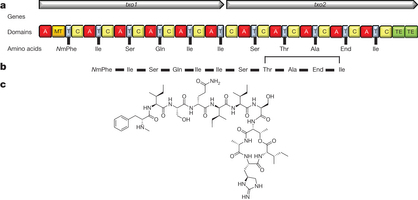
Figure 1: The structure of teixobactin and the predicted biosynthetic gene cluster. a, The two NRPS genes, the catalytic domains they encode, and the amino acids incorporated by the respective modules. Domains: A, adenylation; C, condensation; MT, methylation (of phenylalanine); T, thiolation (carrier); and TE, thioesterase (Ile-Thr ring closure). NmPhe, N-methylated phenylalanine. b, Schematic structure of teixobactin. The N-methylation of the first phenylalanine is catalysed by the methyltransferase domain in module 1. The ring closure between the last isoleucine and threonine is catalysed by the thioesterase domains during molecule off-loading, resulting in teixobactin. c, Teixobactin structure.
-
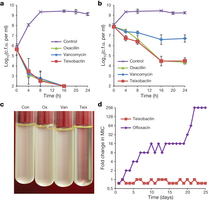
Figure 2: Time-dependent killing of pathogens by teixobactin. a, b, S. aureus were grown to early (a), and late (b) exponential phase and challenged with antibiotics. Data are representative of 3 independent experiments ± s.d. c, Teixobactin treatment resulted in lysis. The figure is representative of 3 independent experiments. d, Resistance acquisition during serial passaging in the presence of sub-MIC levels of antimicrobials. The y axis is the highest concentration the cells grew in during passaging. For ofloxacin, 256 × MIC was the highest concentration tested. The figure is representative of 3 independent experiments.
-
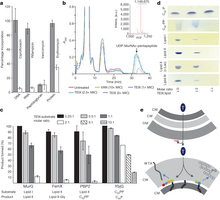
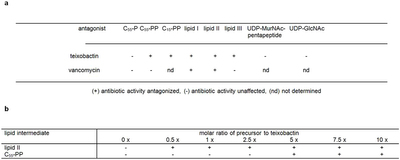
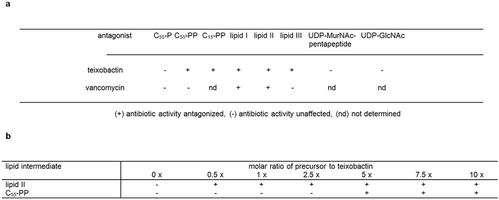
Figure 3: Teixobactin binds to cell wall precursors. a, Impact of teixobactin (TEIX) on macromolecular biosyntheses in S. aureus. Incorporation of 3H-thymidine (DNA), 3H-uridine (RNA), 3H-leucine (protein), and 3H-glucosamine (peptidoglycan) was determined in cells treated with teixobactin at 1 × MIC (grey bars). Ciprofloxacin (8 × MIC), rifampicin (4 × MIC), vancomycin (2 × MIC) and erythromycin (2 × MIC) were used as controls (white bars). Data are means of 4 independent experiments ± s.d. b, Intracellular accumulation of the cell wall precursor UDP-MurNAc-pentapeptide after treatment of S. aureus with teixobactin. Untreated and vancomycin (VAN)-treated (10 × MIC) cells were used as controls. UDP-MurNAc-pentapeptide was identified by mass spectrometry as indicated by the peak at m/z 1,149.675. The experiment is representative of 3 independent experiments. c, The effect of teixobactin on precursor consuming reactions. Experiments were performed in 3 biological replicates and data are presented as mean ± s.d. d, Complex formation of teixobactin with purified cell wall precursors. Binding of teixobactin is indicated by a reduction of the amount of lipid intermediates (visible on the thin-layer chromatogram). The figure is representative of two independent experiments. e, A model of teixobactin targeting and resistance. The teixobactin producer is a Gram-negative bacterium protected from this compound by exporting it across the outer membrane permeability barrier (upper panel). In target Gram-positive organisms lacking an outer membrane, the targets are readily accessible on the outside where teixobactin binds precursors of peptidoglycan (PG) and WTA. CM, cytoplasmic membrane; CW, cell wall; OM, outer membrane; T, teixobactin.
-

Figure 4: Teixobactin is efficacious in three mouse models of infection. a, Single dose treatment (i.v., 1 h post-infection, 6 mice per group) with teixobactin and vancomycin in septicemia protection model using MRSA. Survival is depicted 48 h after infection. b, Single dose (i.v., 2 h post-infection, 4 mice per group) treatment with teixobactin and vancomycin in neutropenic mouse thigh infection model using MRSA. For drug-treated animals, thigh colony-forming units (c.f.u.) were determined at 26 h post-infection. For controls, c.f.u. in thighs were determined at 2 h and 26 h post-infection. c, Two dose treatment, 5 mice per group, with teixobactin (i.v., 24 h and 36 h post-infection) and single dose treatment with amoxicillin (subcutaneous, 24 h post-infection) in immunocompetent lung infection model using S. pneumoniae. Lung c.f.u. were determined at 48 h post-infection. The c.f.u. from each mouse are plotted as individual points and error bars represent the deviation within an experimental group. *P < 0.05, ***P < 0.001 (determined by non-parametric log-rank test).
-
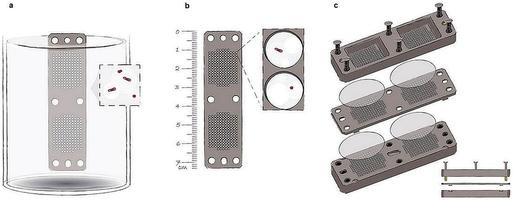
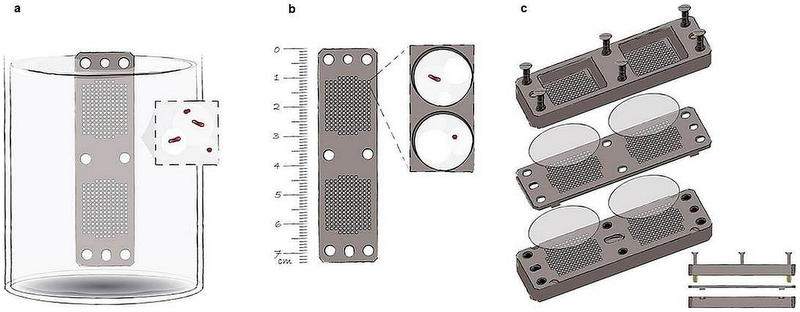
Extended Data Fig. 1: The iChip. a–c, The iChip (a) consists of a central plate (b) which houses growing microorganisms, semi-permeable membranes on each side of the plate, which separate the plate from the environment, and two supporting side panels (c). The central plate and side panels have multiple matching through-holes. When the central plate is dipped into suspension of cells in molten agar, the through-holes capture small volumes of this suspension, which solidify in the form of small agar plugs. Alternatively, molten agar can be dispensed into the chambers. The membranes are attached and the iChip is then placed in soil from which the sample originated.
-
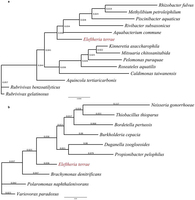
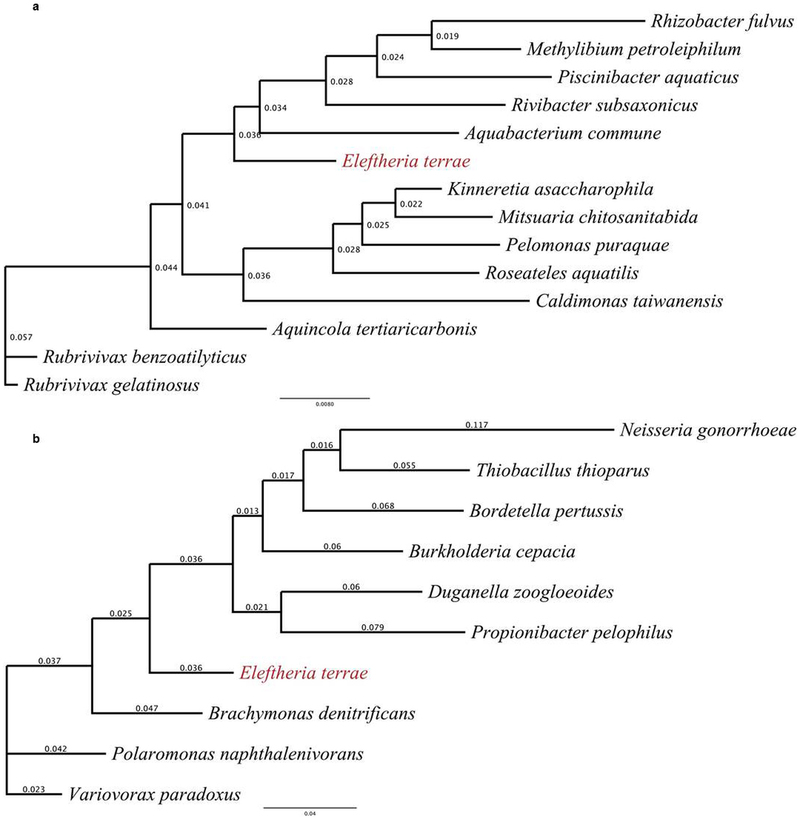
Extended Data Fig. 2: 16S rRNA gene phylogeny of Eleftheria terrae. a, The phylogenetic position of E. terrae within the class β-proteobacteria. The 16S rRNA gene sequences were downloaded from Entrez at NCBI using accession numbers retrieved from peer-reviewed publications. b, The phylogenetic position of E. terrae among its closest known relatives. The sequences were downloaded from NCBI using accession numbers retrieved from the RDP Classifier Database. For both trees, multiple sequence alignments (MSA) were constructed using ClustalW2, implementing a default Cost Matrix, the Neighbour-Joining (NJ) clustering algorithm, as well as optimized gap penalties. Resulting alignments were manually curated and phylogenetic trees were constructed leveraging PhyML 3.0 with a TN93 substitution model and 500 Bootstrap iterations of branch support. Topology search optimization was conducted using the Subtree–Pruning–Regrafting (SPR) algorithm with an estimated Transition–Transversion ratio and gamma distribution parameters as well as fixed proportions of invariable sites.
-

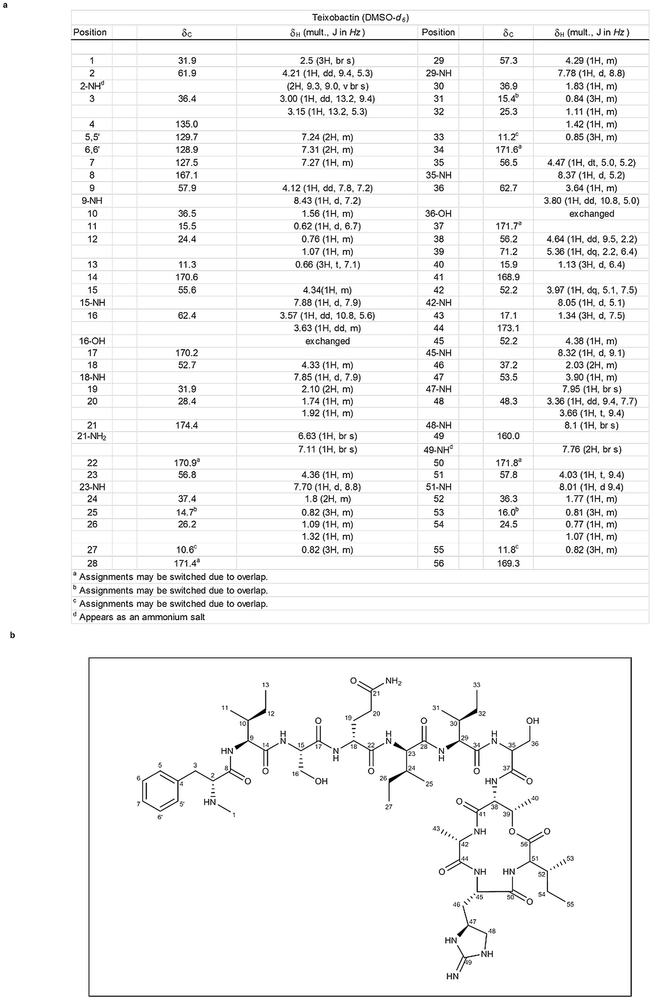
Extended Data Fig. 3: NMR assignment of teixobactin. a, 13C-NMR of teixobactin (125 mHz, δ in p.p.m.). b, Structure of teixobactin with the NMR assignments.
-

Extended Data Fig. 4: NMR spectra of teixobactin. a, 13C NMR spectrum of teixobactin. b, 1H NMR spectrum. c, HMBC NMR spectrum. d, HSQC NMR spectrum. e, COSY NMR spectrum.
-
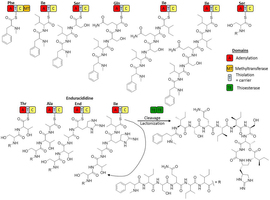
Extended Data Fig. 5: Hypothetical biosynthesis pathway of teixobactin. The eleven modules of the non-ribosomal peptide synthetases Txo1 and Txo2 are depicted with the growing chain attached. Each module is responsible for the incorporation of one specific amino acid in the nascent peptide chain. The N-methylation of the first amino acid phenylalanine is catalysed by the methyltransferase domain in module 1. The ring closure (marked by a dashed arrow) between the last isoleucine and threonine is catalysed by the thioesterase domains during molecule off-loading, resulting in teixobactin.
-
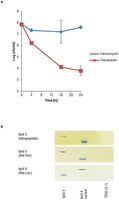
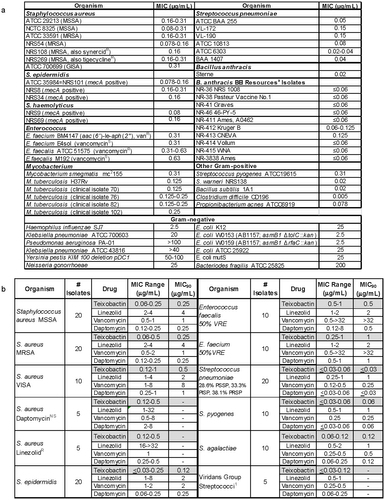
Extended Data Fig. 6: Teixobactin activity against vancomycin-resistant strains. a, Vancomycin intermediate S. aureus (VISA) were grown to late exponential phase and challenged with vancomycin or teixobactin. Cell numbers were determined by plating for colony counts. Data are representative of 3 independent experiments ± s.d. b, Complex formation of teixobactin with cell wall precursor variants as formed by vancomycin-resistant strains. Purified lipid intermediates with altered stem peptides were incubated with teixobactin at a molar ratio of 2:1 (TEIX:lipid II variant). Reaction mixtures were extracted with BuOH/PyrAc and binding of teixobactin to lipid II variants is indicated by its absence on the thin-layer chromatogram. Migration behaviour of unmodified lipid II is used for comparison. The figure is representative of 3 independent experiments.
-
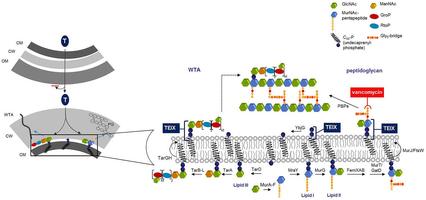
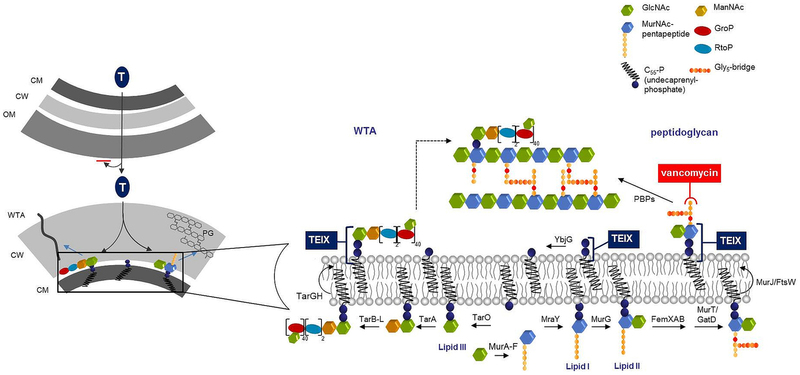
Extended Data Fig. 7: Model for the mechanism of action of teixobactin. Inhibition of cell wall synthesis by teixobactin. Lipid II, precursor of peptidoglycan, is synthesized in the cytoplasm and flipped to the surface of the inner membrane by MurJ48 or FtsW49. Lipid III, a precursor of wall teichoic acid (WTA), is similarly formed inside the cell and WTA lipid-bound precursors are translocated across the cytoplasmic membrane by the ABC-transporter TarGH50. Teixobactin (TEIX) forms a stoichiometric complex with cell wall precursors, lipid II and lipid III. Abduction of these building blocks simultaneously interrupts peptidoglycan (right), WTA (left) biosynthesis as well as precursor recycling. Binding to multiple targets within the cell wall pathways obstructs the formation of a functional cell envelope. Left panel, teixobactin targeting and resistance. The producer of teixobactin is a Gram-negative bacterium which is protected from this compound by exporting it outside of its outer membrane permeability barrier. The target Gram-positive organisms do not have an outer membrane. CM, cytoplasmic membrane; CW, cell wall; OM, outer membrane; LTA, lipoteichoic acid; WTA, wall teichoic acid.
-
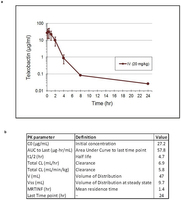
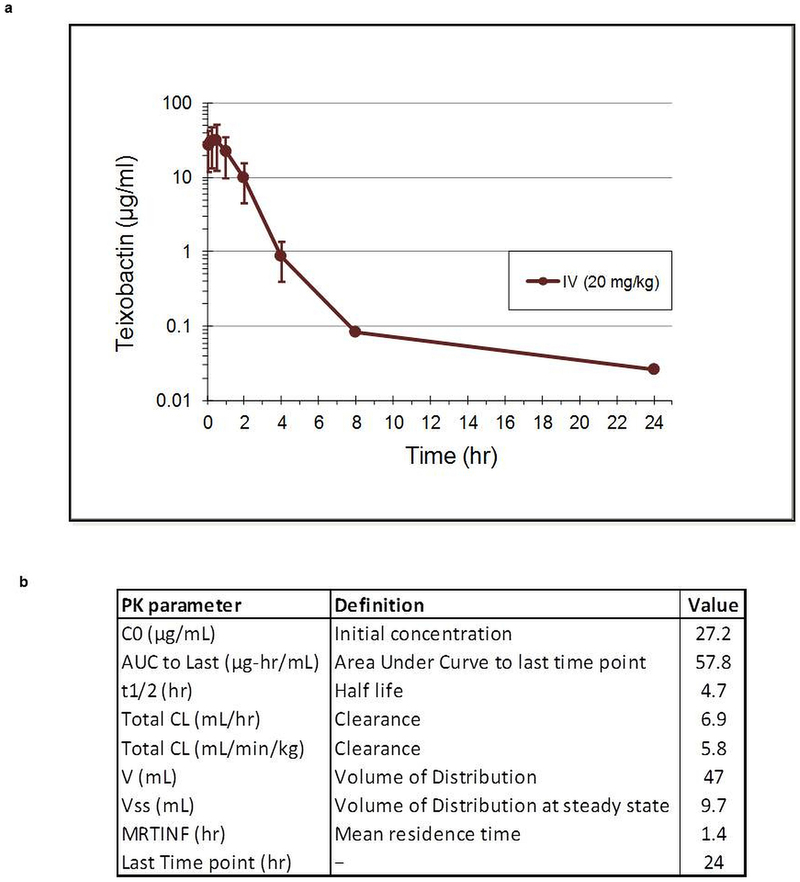
Extended Data Fig. 8: Pharmacokinetic analysis of teixobactin. a, The mean plasma concentrations of teixobactin after a single i.v. injection of 20 mg per kg teixobactin (3 mice per time point). Data are the mean of plasma concentration, and error bars represent the standard deviation from 3 animals in each time point. b, Pharmacokinetic parameters of teixobactin calculated with a non-compartmental analysis model based on WinNonlin.

Read the full article
Additional access options:
- British Library Document Supply Centre
- You can also request this document from your local library through inter-library loan services.